October 7, 2002
The countermanding paradigm, which includes both a task design and a specific theoretical construct, was developed to investigate the control of action (e.g. Vince 1948; Lappin and Eriksen 1966; DeJong et al., 1990, 1995; reviewed by Logan and Cowan 1984; Logan 1994). A subject's ability to control voluntarily the production of movements is evaluated in a reaction time task by infrequently presenting an imperative stop-signal. The subject is instructed to withhold the impending movement if the stop-signal occurs.
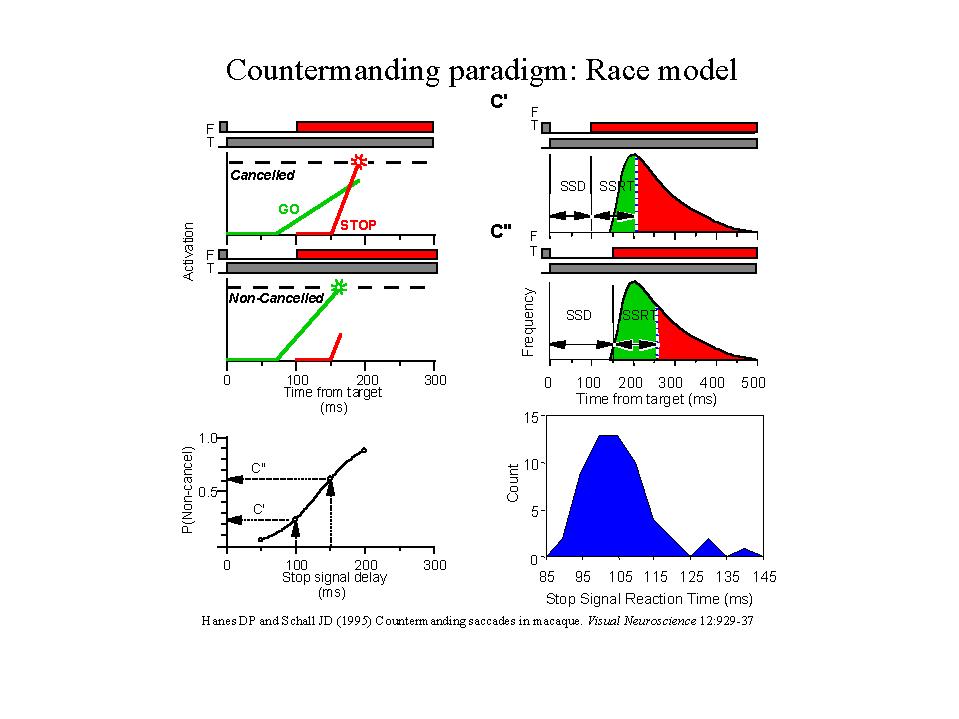
Performance in the countermanding task is probabilistic. On a given trial one can predict only to a certain extent whether the subject will be able to inhibit a planned movement. The probability of inhibiting a movement decreases as the delay between the signal to initiate the movement and the signal to inhibit the movement, called the stop-signal, increases. This unpredictability arises because response time is fundamentally stochastic, varying unpredictably across trials. In principle one can see that movements generated with short latencies would occur even if the stop-signal were presented because such short-latency movements would be initiated before the stop-signal could influence the system. Likewise, movements generated with long latencies would be inhibited if a stop-signal was presented because their reaction times allow enough time for the stop-signal to influence the system thereby canceling the planned movement. These relationships permit an experimental comparison between trials in which a stop-signal was presented and movement production was successfully inhibited and trials with movements that were made but would have been inhibited had the stop-signal been presented (the trials with the long reaction times). By comparing the neural activity in these different trial types one can investigate the neural mechanisms underlying the gaze holding and gaze shifting processes.
The data obtained in the countermanding task are the inhibition function and the distribution of reaction times in NO STOP SIGNAL trials. The inhibition function plots the probability of the monkey generating a movement to the target (NON-CANCELLED trials) as a function of stop-signal delay. The inhibition functions show that following short stop-signal delays, the monkeys successfully withheld movements to the target. But as the stop-signal delay increased, the monkeys increasingly failed to withhold the movement. Note that the probability of NON-CANCELLED trials is equal to 1.0 minus the probability of CANCELLED trials.
A critical value used in these investigations is the length of time required to cancel the movement being programmed. This duration, known as the stop-signal reaction time (SSRT), is a measure that is not directly available in the behavioral data. However, the application of a race model provides a means of estimating the duration of this covert inhibitory process (Logan and Cowan 1984; Logan 1994). The model consists of a race between a GO process and a STOP process. The GO process prepares and generates the movement following the presentation of the target. When the stop-signal is not given, only the GO process is active (NO STOP SIGNAL trials). Thus, the distribution of movement latencies obtained in NO STOP SIGNAL trials is the distribution of finish times of the GO process. If the stop-signal is given, then while the GO process proceeds, the STOP process is invoked. If the STOP process finishes before the GO process, then the movement will not be produced resulting in a CANCELLED trial. Alternatively, if the GO process finishes before the STOP process, then the movement will be generated resulting in a NON-CANCELLED trial.
The increasing inhibition function arises because increasing the stop-signal delay postpones the onset of the STOP process, thus increasing the probability that the GO process will finish before the STOP process finishes. Following shorter stop signal delays the STOP process finishes more often before the GO process, resulting in a lower fraction of NON-CANCELLED trials. Following a longer stop signal delay the STOP process finishes less often before the GO process, resulting in a higher fraction of NON-CANCELLED trials.
Two methods of estimation of the SSRT have been developed (Logan and Cowan, 1984; Logan, 1994). The first method of estimating the SSRT assumes that it is a random variable. The mean SSRT is equal to the difference between the mean reaction time during NO STOP SIGNAL trials and the mean value of the inhibition function. The mean of the inhibition function is determined by treating the inhibition function as a cumulative distribution and converting it to a probability density function. If the inhibition function ranges from a probability of 0 to 1, then the mean is the difference between the probability of responding at the ith stop signal delay minus the probability of responding at the i-1th stop signal delay multiplied by the ith stop signal delay, summed over all stop signal delays (Logan and Cowan, 1984).
A second method of calculating the SSRT provides an estimate at each
stop-signal delay by making the convenient but nonessential assumption
that the SSRT is constant. Although this assumption seems unwarranted because
it is implausible that a physiological process would take a constant amount
of time to execute, its violation does not substantially change the outcome
of this analysis (Logan and Cowan, 1984; DeJong et al., 1990; Band, 1997).
By this method the SSRT is estimated by integrating the NO STOP SIGNAL
movement latency distribution, beginning at the time of target presentation,
until the integral equals the proportion of NON-CANCELLED trials at that
stop-signal delay. The movement latency at the limit of the integral represents
the finish line of the stop process. In other words that time value represents
the longest movement latency at which the GO process finished before the
STOP process inhibited the movement. Thus, the time between the appearance
of the stop-signal and this finish line is the SSRT at this stop-signal
delay. In practice, the SSRT is determined by rank ordering the NO STOP
SIGNAL movement latencies. The ith movement latency is then
chosen, where i is determined by multiplying the probability
of a NON-CANCELLED trial at a given stop-signal delay times the total number
of NO STOP SIGNAL trials. The SSRT is the difference between the ith
movement latency and the stop-signal delay.
References
DeJong, R., Coles, M.G.H., Logan, G.D. AND Gratton, G. In search of the point of no return: The control of response processes. J. Exp. Psychol.: Human Percept. Perform. 16:164-182, 1990.
DeJong, R., Coles, M.G.H., AND Logan, G.D. Strategies and mechanisms in nonselective and selective inhibitory motor control. J. Exp. Psychol.: Human Percept. Perform. 21:498-511, 1995.
Hanes, D.P., AND Schall, J.D. Countermanding saccades in macaque. Vis. Neurosci 12: 929-937, 1995.
Lappin, J.S. AND Eriksen, C.W. Use of a delayed signal to stop a visual reaction-time response. J. Exp. Psych. 72: 805-811, 1966.
Logan, G.D. On the ability to inhibit thought and action: A users' guide to the stop signal paradigm. In: Inhibitory Processes in Attention, Memory and Language, edited by D. Dagenbach, T.H. Carr. San Diego: Academic Press. 1994, p. 189-239.
Logan, G.D., AND Cowan, W.B. On the ability to inhibit thought and action: A theory of an act of control. Psych. Rev. 91:295-327, 1984.
Vince, M.A. The intermittency of control movements and the psychological
refractory period.
Brit. J. Psychol. 38:149-157, 1948.
{kind=link}
{kind=link}
{kind=link}
{kind=link}